Фотосинтез и его регуляция
Все живое на Земле зависит от фотосинтеза. Фотосинтез занимает центральное место в круговороте вещества и энергии на Земле, делая энергию и углерод доступными для организмов и обеспечивая выделение кислорода в атмосферу. Познание механизмов утилизации света в фотосинтезе играет первостепенную роль для управления этим процессом и имеет большое общебиологическое значение для понимания основных принципов биоэнергетики.
В настоящее время в основном установлены состав и организация фотосинтетического аппарата и механизмы отдельных стадий, начиная от поглощения квантов света и кончая образованием углеводов. Вместе с тем, вопрос о регуляции процессов фотосинтеза, обеспечивающей оптимальное функционирование фотосинтетического аппарата в целом, еще далек от разрешения.
Структурно-функциональная саморегуляция является важнейшим свойством всех биологических систем. Регуляторные механизмы определяют устойчивость системы, ее способность адаптироваться к изменяющимся условиям внешней среды (температура, газовый и химический состав, освещенность и т.д.).
Флуоресценция хлорофилла является пока единственным показателем, который позволяет исследовать в живых объектах протекание фотохимических реакций, связанных с работой так называемой фотосистемы 2 (ФС2) высших растений – системы, наиболее чувствительной к факторам внешней среды. В наибольшей мере регуляторные механизмы фотосинтеза проявляются в различных индукционных явлениях и, в частности, в явлении так называемой медленной индукции флуоресценции (МИФ) фотосинтезирующих объектов. МИФ весьма чувствительна к состоянию фотосинтетического аппарата, что, наряду с простой техникой эксперимента, обусловливает перспективность ее изучения.
В последние годы для характеристики функциональной активности фотосинтетического аппарата используют большое число флуоресцентных показателей, связанных с МИФ. Это обусловлено тем, что их изменения коррелируют с изменением интенсивности фотосинтеза. Результаты таких исследований способствуют более глубокому пониманию регуляторных механизмов, обеспечивающих эффективное преобразование энергии в первичных и последующих стадиях фотосинтеза. Кроме того, они могут быть использованы как для построения различных математических моделей фотосинтеза, так и при проведении экологического мониторинга растительных объектов.
Структурно-функциональная организация фотосинтетического аппарата высших растений
Фотосинтез осуществляют высшие растения, водоросли и некоторые бактерии. Этот процесс заключается в трансформации поглощенной световой энергии в химическую энергию органических соединений. Общее уравнение фотосинтеза:
![]()
![]()
Суммарный процесс фотосинтеза высших растений можно разделить на две взаимосвязанные стадии: световую и темновую. Световая стадия включает поглощение света пигментами, которые при этом переходят в возбужденное состояние, миграцию энергии электронного возбуждения к реакционным центрам (РЦ), разделение зарядов и трансформацию энергии в РЦ, перенос электронов по электронтранспортной цепи (ЭТЦ) и сопряженные с ним процессы, приводящие к образованию первичных стабильных продуктов: НАДФ.Н и АТФ. Темновая стадия включает ассимиляцию СО2 и образование конечных продуктов в реакциях цикла Кальвина, которые протекают с использованием НАДФ.Н и АТФ.
В зеленых растениях все световые и часть темновых стадий фотосинтеза протекают в субклеточных частицах – хлоропластах.
Хлоропласты высших растений на срезе обычно имеют двояковыпуклую форму, а при взгляде сверху выглядят округлыми. Диаметр хлоропластов составляет от 3 до 10 мкм. Их внутреннее содержимое состоит из стромы (матрикса), в которой протекают темновые стадии фотосинтеза, и ламеллярной системы, образованной мембранами тилакоидов, – замкнутых дискообразных структур диаметром около 500 нм, состоящих из близко расположенных мембран толщиной 90 Aо (рис. 1,А). В мембранах тилакоидов происходят световые стадии фотосинтеза.
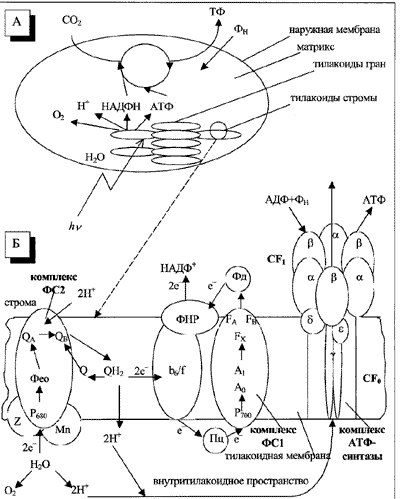
Рис. 1. Схематическое изображение хлоропласта (А) и первичных процессов фотосинтеза (Б).
Обозначения см. в подписи к рис. 2
Внутри одного хлоропласта содержится обычно около 1 тыс. тилакоидов. Различают тилакоиды гран (граны – стопки тилакоидов) и тилакоиды стромы, выходящие за пределы гран и являющиеся продолжением тилакоидов гран. На один тилакоидный диск приходится в среднем 200 ЭТЦ и 105–106 молекул пигментов (хлорофилл а (Хл а), хлорофилл b (Хл b), каротиноиды).
Большинство экспериментальных данных о первичных процессах фотосинтеза у высших растений хорошо укладывается в так называемую Z-схему фотосинтеза (рис. 2). По этой схеме перенос электрона от Н2О к НАДФ+ при поглощении двух квантов света последовательно осуществляется двумя фотосистемами (ФС): коротковолновой ФС2, поглощающей свет с длиной волны =< 690 нм, и длинноволновой ФС1, поглощающей свет с длиной волны =< 710 нм.
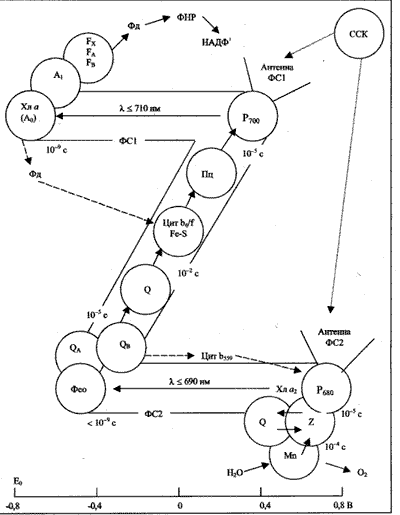
Рис. 2. Схема последовательного переноса электрона от воды к НАДФ в световой стадии фотосинтеза. ССК – светособирающий комплекс; A0 – первичный акцептор электрона ФС1 – молекула хлорофилла а; вторичные акцепторы – молекулы филлохинона (А1); три переносчика белковой природы – ферредоксины FX, FA, FB; Фео – первичный акцептор ФС2 – феофитин; вторичные акцепторы ФС2 – молекулы пластохинона QA и QB; Цит – цитохромы; Пц – пластоцианин; Fe-S – железо-серный центр Риске; Фд – ферредоксин; ФНР – Фд-НАДФ-редуктаза; Z – промежуточный переносчик в системе разложения воды; штриховые линии – циклические потоки электронов. Внизу приведена шкала окислительно-восстановительных потенциалов
Каждая ФС представляет собой обособленную структуру ламеллярной мембраны. Эта структура состоит из следующих компонентов: матрицы светособирающих пигментов; фотохимически активного хлорофилла, принимающего участие в разделении зарядов и входящего в состав РЦ; нескольких переносчиков электронов белковой и небелковой природы. Комплексы ФС2 локализованы в основном в уплотненных областях тилакоидов гран, а комплексы ФС1 находятся преимущественно в стромальных участках тилакоида и в неуплотненных областях тилакоидов гран.
Энергия квантов света, попадающих в фотосинтетический аппарат (ФА), может передаваться на РЦ несколькими путями. Во-первых, каждая ФС имеет собственную (коровую) хлорофилльную антенну (20–40 молекул Хл а), эффективно захватывающую фотоны и передающую энергию возбуждения на РЦ. Во-вторых, энергия возбуждения может поступать в ФС из периферической антенны, образованной светособирающим комплексом (ССК). В ССК сосредоточена примерно половина имеющегося в хлоропластах Хл a и практически весь Хл b. В настоящее время установлены структуры ФС1 (с разрешением 4,5 Aо), ФС2 (с разрешением 8 Aо) и ССК (с разрешением 3,4 Aо).
Хл a является компонентом как коровых комплексов, так и периферической антенны ФС1 и ФС2, тогда как Хл b преимущественно является компонентом периферической антенны обеих фотосистем. В связи с этим изменение отношения Хл а/Хл b указывает на изменение соотношения между комплексами реакционных центров фотосистем и ССК.
Часть ССК, связанная с ФС1 (ССК1), находится в тилакоидах стромы, а часть, связанная с ФС2 (ССК2), – в тилакоидах гран (70–90% всех ССК). Основной структурной единицей этого комплекса является белок с молекулярной массой 25–30 кДа, с которым связаны 7–8 молекул Хл а, 5–6 молекул Хл b и каротиноиды: лютеин, неоксантин и виолаксантин. Помимо улавливания света, каротиноиды выполняют важную защитную функцию, принимая на себя избыток энергии от возбужденных молекул хлорофилла и предотвращая образование активных форм кислорода.
Часть комплексов ССК2 подвижна, она может работать либо на ФС1, либо на ФС2, т.к. способна диффундировать из стромальной области в гранальную. Подвижные ССК2 играют важную роль в распределении энергии квантов света между двумя ФС. Оно связано с обратимым фосфорилированием белка ССК, которое влияет на соотношение между нециклическим и циклическим электронными потоками, обеспечивая требуемое соотношение между количествами НАДФ.Н и АТФ (см. ниже).
Молекулы хлорофилла и вспомогательных пигментов поглощают солнечный свет и переходят в возбужденное состояние. Энергия возбуждения мигрирует по пигментной матрице и попадает в конце концов на РЦ: Р680 в ФС2 и Р700 в ФС1. Здесь происходит разделение положительных и отрицательных зарядов, после чего электрон переносится на первичные акцепторы. На каждую ЭТЦ приходится 300–400 молекул пигментов, что обусловливает частое срабатывание РЦ даже при небольшой интенсивности света. Резонансный перенос энергии между молекулами-светосборщиками и захват ее в РЦ происходят за время около 10–12 с, однако разделенные заряды в РЦ стабилизируются на время около 10–2 с, которого достаточно для эффективного переноса электронов на стадиях, ограниченных диффузией.
Окисленный РЦ ФС2 (P+680) имеет окислительно-восстановительный потенциал +0,81 В и может окислить молекулу воды с помощью специальной ферментативной системы, входящей в состав кислород-выделяющего комплекса. Для функционирования этой системы необходимы ионы Mn:
![]()
![]()
Первичный акцептор ФС1 имеет окислительно-восстановительный потенциал –0,62 В и может восстанавливать конечный акцептор электронов НАДФ+ через ферредоксин-НАДФ— редуктазу.
Между комплексами ФС1 и ФС2 электроны переносятся по градиенту окислительно-восстановительного потенциала через ЭТЦ. ЭТЦ включает подвижный переносчик пластохинон, цитохромный комплекс b6–f, в состав которого входят цитохром f и два цитохрома b6, а также медьсодержащий белок пластоцианин (Пц), являющийся связующим звеном между комплексом b6–f и Р700 (рис. 1Б, 2).
Описанная совокупность процессов переноса электронов называется нециклическим транспортом. Его лимитирующей стадией является окисление пластохинона комплексом b6–f (t0,5 ~ 15–20 мс и более).
При замедлении или прекращении окисления НАДФ.Н в ФС1 осуществляется циклический электронный транспорт (рис. 2). Таким образом, в зависимости от количества НАДФ+ в системе реализуется либо нециклический транспорт электронов с участием обеих ФС, либо циклический, при котором работает лишь ФС1. Локализация большей части ФС2 в гранальной области обеспечивает основной нециклический поток электронов, восстанавливающий молекулы пластохинона, которые затем диффундируют в стромальную область, отдавая электроны ФС1.
Другая часть ФС2, локализованная в стромальной области, обеспечивает более слабый поток, взаимодействующий с интенсивным циклическим потоком ФС1 – источником энергии для синтеза АТФ. Есть данные, что ферредоксин-НАДФ-редуктаза образует единый комплекс с цитохромным комплексом b6–f. Благодаря этому замыкается циклический путь электронов, включающий ФС1 и комплекс b6-f.
Как было отмечено выше, распределение электронов по нециклическому и циклическому пути имеет важное значение для достижения оптимального соотношения между количествами синтезируемого АТФ и восстановленного НАДФ.Н. Считается, что определенную регуляторную роль играет также циклический электронный транспорт вокруг ФС2.
Процессы электронного и протонного транспорта приводят в действие АТФ-синтазу – фермент, расположенный в стромальной области тилакоидов и катализирующий синтез молекулы АТФ. На рис. 1,Б схематично представлено пространственное расположение комплекса, образующего АТФ.
АТФ-синтаза представляет собой белковый ансамбль, который состоит из двух крупных белковых фрагментов: локализованного в мембране гидрофобного фактора сопряжения CF0 и выступающего наружу (в сторону стромы) белкового комплекса (фактор сопряжения CF1). Реакции синтеза АТФ происходят поочередно в трех b-субъединицах фактора сопряжения CF1, которые вместе с тремя гомологичными им a-субъединицами образуют шарообразную структуру.
В нативной ЭТЦ формально можно выделить два места сопряжения электронного транспорта и фосфорилирования, связанные с выделением протонов во внутритилакоидное пространство: на донорной стороне ФС2 и между двумя ФС (рис. 1,Б). Работу АТФ-синтазы обеспечивает обратный поток протонов через комплекс CF0-CF1, направленный из тилакоидов в строму. При этом мембранный фрагмент CF0 выполняет роль протон-проводящего канала. В последние годы было установлено, что переносящий протоны АТФ-синтазный комплекс работает как настоящая молекулярная машина с вращающимся ротором.
Существенно, что скорость переноса электронов зависит от соотношения между количеством субстратов и продуктов реакции синтеза АТФ:
![]()
![]()
При избытке АДФ и ФН протоны активно проходят через АТФ-синтазу, сильного закисления внутритилакоидного пространства не происходит и скорость электронного транспорта поддерживается на высоком уровне. По мере истощения АДФ или ФН канал быстрого выхода протонов наружу через АТФ-синтазу закрывается, значение внутритилакоидного pH понижается и перенос электронов между ФС (окисление пластохинона) замедляется. Когда АДФ и ФН появляются вновь (например, за счет гидролиза АТФ в последующих биохимических реакциях), протоны снова начинают «прокачиваться» через АТФ-синтазу, их концентрация внутри тилакоидов уменьшается, скорость электронного транспорта возрастает и т.д.
АТФ и НАДФ.Н, образованные в световой стадии фотосинтеза, используются далее в темновой стадии фиксации СО2. Ферменты, катализирующие отдельные стадии цикла, растворимы в воде и локализованы в строме хлоропластов.
Единственным путем, посредством которого СО2 превращается в фосфаты сахаров, у всех фотоавтотрофных растений является восстановительный пентозофосфатный цикл (ВПФ-цикл), или цикл Кальвина-Бенсона-Бассэма, для которого требуется соотношение АТФ/НАДФ.Н, равное 3:2 (для связывания одной молекулы СО2 затрачиваются 3 молекулы АТФ и 2 молекулы НАДФ.Н). Суммарное уравнение реакций цикла имеет вид:
![]()
![]()
При нециклическом переносе на 2e образуется 2АТФ и 2НАДФ.Н, т.е. нециклический транспорт электронов не может полностью удовлетворить потребность ВПФ-цикла в АТФ. Для получения дополнительного АТФ используется циклический электронный транспорт в ФС1, упоминавшийся выше.
Таким образом, структурная организация фотосинтетического аппарата растений в настоящее время изучена достаточно хорошо. Наибольшее внимание исследователей привлекают те регуляторные механизмы, которые обеспечивают оптимальное функционирование этого аппарата и наиболее эффективное протекание всех фотосинтетических процессов. Многие из этих механизмов находят отражение в индукционных изменениях флуоресценции фотосинтезирующих объектов и, в частности, в явлении медленной индукции флуоресценции (МИФ).
Современные представления о медленной индукции флуоресценции фотосинтезирующих объектов
При освещении растительной клетки достаточно коротковолновым светом (например, синим), наблюдается ее свечение – испускание света с большей длиной волны (например, красного), называемое флуоресценцией.
Источником флуоресценции в растительной клетке являются светособирающие молекулы пигментов, причем подавляющая часть флуоресценции в нормальных условиях обусловлена Хл а ФС2.
Квантовый выход флуоресценции (отношение числа испущенных квантов к числу поглощенных) равен:
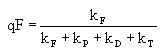
где kF – константа скорости флуоресценции; kP – константа скорости фотохимической реакции; kD – константа скорости тепловой дезактивации; kT – константа скорости переноса энергии возбуждения к нефлуоресцирующим пигментам.
Квантовый выход флуоресценции Хл а in vivo составляет 2–8% со средним временем жизни около 300 пс (для «открытых» РЦ) и около 1,6 нс (для «закрытых» РЦ). Вклад ФС1 в сигнал флуоресценции при комнатной температуре не превышает 20%, а в изолированной ФС1 он составляет менее 1% из-за короткого времени жизни возбужденных состояний (t~10–10–10–9 с). При комнатной температуре флуоресценция определяется количеством светящихся молекул Хл а в ССК и ФС2, в то время как вклад хлорофилла ФС1 становится заметным в случае проявления эффекта «спилловера» – передачи энергии возбуждения из ФС2 в ФС1.
Источник: И.Б. ПОЛЯКОВА. ОБЩАЯ БИОЛОГИЯ. Фотосинтез и его регуляция.