Фотосинтез — синтез органических веществ из углекислого газа и воды с обязательным использованием энергии света:
6СО2 + 6Н2О + Qсвета → С6Н12О6 + 6О2.
У высших растений органом фотосинтеза является лист, органоидами фотосинтеза — хлоропласты. В мембраны тилакоидов хлоропластов встроены фотосинтетические пигменты: хлорофиллы и каротиноиды. Существует несколько разных типов хлорофилла (a, b, c, d), главным является хлорофилл a. В молекуле хлорофилла можно выделить порфириновую «головку» с атомом магния в центре и фитольный «хвост». Порфириновая «головка» представляет собой плоскую структуру, является гидрофильной и поэтому лежит на той поверхности мембраны, которая обращена к водной среде стромы. Фитольный «хвост» — гидрофобный и за счет этого удерживает молекулу хлорофилла в мембране.
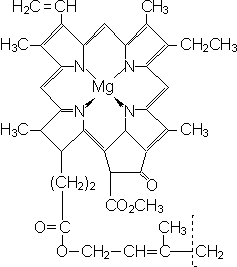
Хлорофиллы поглощают красный и сине-фиолетовый свет, отражают зеленый и поэтому придают растениям характерную зеленую окраску. Молекулы хлорофилла в мембранах тилакоидов организованы в фотосистемы. У растений и синезеленых водорослей имеются фотосистема-1 и фотосистема-2, у фотосинтезирующих бактерий — фотосистема-1. Только фотосистема-2 может разлагать воду с выделением кислорода и отбирать электроны у водорода воды.
Фотосинтез — сложный многоступенчатый процесс; реакции фотосинтеза подразделяют на две группы: реакции световой фазы и реакции темновой фазы.
Световая фаза
Эта фаза происходит только в присутствии света в мембранах тилакоидов при участии хлорофилла, белков-переносчиков электронов и фермента — АТФ-синтетазы. Под действием кванта света электроны хлорофилла возбуждаются, покидают молекулу и попадают на внешнюю сторону мембраны тилакоида, которая в итоге заряжается отрицательно. Окисленные молекулы хлорофилла восстанавливаются, отбирая электроны у воды, находящейся во внутритилакоидном пространстве. Это приводит к распаду или фотолизу воды:
Н2О + Qсвета → Н+ + ОН—.
Ионы гидроксила отдают свои электроны, превращаясь в реакционноспособные радикалы •ОН:
ОН— → •ОН + е—.
Радикалы •ОН объединяются, образуя воду и свободный кислород:
4НО• → 2Н2О + О2.
Кислород при этом удаляется во внешнюю среду, а протоны накапливаются внутри тилакоида в «протонном резервуаре». В результате мембрана тилакоида с одной стороны за счет Н+ заряжается положительно, с другой за счет электронов — отрицательно. Когда разность потенциалов между наружной и внутренней сторонами мембраны тилакоида достигает 200 мВ, протоны проталкиваются через каналы АТФ-синтетазы и происходит фосфорилирование АДФ до АТФ; атомарный водород идет на восстановление специфического переносчика НАДФ+ (никотинамидадениндинуклеотидфосфат) до НАДФ·Н2:
2Н+ + 2е— + НАДФ → НАДФ·Н2.
Таким образом, в световую фазу происходит фотолиз воды, который сопровождается тремя важнейшими процессами: 1) синтезом АТФ; 2) образованием НАДФ·Н2; 3) образованием кислорода. Кислород диффундирует в атмосферу, АТФ и НАДФ·Н2транспортируются в строму хлоропласта и участвуют в процессах темновой фазы.
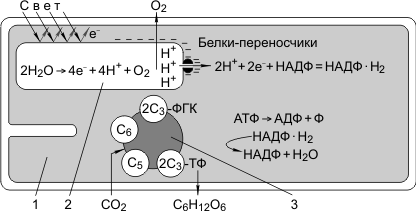
1 — строма хлоропласта; 2 — тилакоид граны.
Темновая фаза
Эта фаза протекает в строме хлоропласта. Для ее реакций не нужна энергия света, поэтому они происходят не только на свету, но и в темноте. Реакции темновой фазы представляют собой цепочку последовательных преобразований углекислого газа (поступает из воздуха), приводящую к образованию глюкозы и других органических веществ.
Первая реакция в этой цепочке — фиксация углекислого газа; акцептором углекислого газа является пятиуглеродный сахар рибулозобифосфат (РиБФ); катализирует реакцию фермент рибулозобифосфат-карбоксилаза (РиБФ-карбоксилаза). В результате карбоксилирования рибулозобисфосфата образуется неустойчивое шестиуглеродное соединение, которое сразу же распадается на две молекулы фосфоглицериновой кислоты (ФГК). Затем происходит цикл реакций, в которых через ряд промежуточных продуктов фосфоглицериновая кислота преобразуется в глюкозу. В этих реакциях используются энергии АТФ и НАДФ·Н2, образованных в световую фазу; цикл этих реакций получил название «цикл Кальвина»:
6СО2 + 24Н+ + АТФ → С6Н12О6 + 6Н2О.
Кроме глюкозы, в процессе фотосинтеза образуются другие мономеры сложных органических соединений — аминокислоты, глицерин и жирные кислоты, нуклеотиды. В настоящее время различают два типа фотосинтеза: С3— и С4-фотосинтез.
С3-фотосинтез
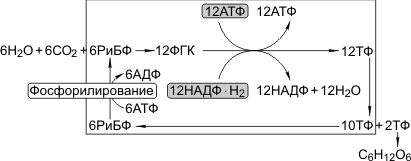
Это тип фотосинтеза, при котором первым продуктом являются трехуглеродные (С3) соединения. С3-фотосинтез был открыт раньше С4-фотосинтеза (М. Кальвин). Именно С3-фотосинтез описан выше, в рубрике «Темновая фаза». Характерные особенности С3-фотосинтеза: 1) акцептором углекислого газа является РиБФ, 2) реакцию карбоксилирования РиБФ катализирует РиБФ-карбоксилаза, 3) в результате карбоксилирования РиБФ образуется шестиуглеродное соединение, которое распадается на две ФГК. ФГК восстанавливается до триозофосфатов (ТФ). Часть ТФ идет на регенерацию РиБФ, часть превращается в глюкозу.
Фотодыхание
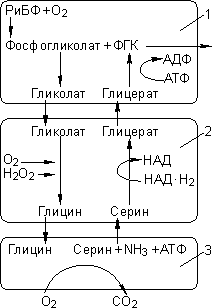
Фотодыхание:
1 — хлоропласт; 2 — пероксисома; 3 — митохондрия.
Это светозависимое поглощение кислорода и выделение углекислого газа. Еще в начале прошлого века было установлено, что кислород подавляет фотосинтез. Как оказалось, для РиБФ-карбоксилазы субстратом может быть не только углекислый газ, но и кислород:
О2 + РиБФ → фосфогликолат (2С) + ФГК (3С).
Фермент при этом называется РиБФ-оксигеназой. Кислород является конкурентным ингибитором фиксации углекислого газа. Фосфатная группа отщепляется, и фосфогликолат становится гликолатом, который растение должно утилизировать. Он поступает в пероксисомы, где окисляется до глицина. Глицин поступает в митохондрии, где окисляется до серина, при этом происходит потеря уже фиксированного углерода в виде СО2. В итоге две молекулы гликолата (2С + 2С) превращаются в одну ФГК (3С) и СО2. Фотодыхание приводит к понижению урожайности С3-растений на 30–40% (С3-растения — растения, для которых характерен С3-фотосинтез).
С4-фотосинтез
С4-фотосинтез — фотосинтез, при котором первым продуктом являются четырехуглеродные (С4) соединения. В 1965 году было установлено, что у некоторых растений (сахарный тростник, кукуруза, сорго, просо) первыми продуктами фотосинтеза являются четырехуглеродные кислоты. Такие растения назвали С4-растениями. В 1966 году австралийские ученые Хэтч и Слэк показали, что у С4-растений практически отсутствует фотодыхание и они гораздо эффективнее поглощают углекислый газ. Путь превращений углерода в С4-растениях стали называть путем Хэтча-Слэка.
Для С4-растений характерно особое анатомическое строение листа. Все проводящие пучки окружены двойным слоем клеток: наружный — клетки мезофилла, внутренний — клетки обкладки. Углекислый газ фиксируется в цитоплазме клеток мезофилла, акцептор — фосфоенолпируват (ФЕП, 3С), в результате карбоксилирования ФЕП образуется оксалоацетат (4С). Процесс катализируется ФЕП-карбоксилазой. В отличие от РиБФ-карбоксилазы ФЕП-карбоксилаза обладает большим сродством к СО2 и, самое главное, не взаимодействует с О2. В хлоропластах мезофилла много гран, где активно идут реакции световой фазы. В хлоропластах клеток обкладки идут реакции темновой фазы.
Оксалоацетат (4С) превращается в малат, который через плазмодесмы транспортируется в клетки обкладки. Здесь он декарбоксилируется и дегидрируется с образованием пирувата, СО2 и НАДФ·Н2.
Пируват возвращается в клетки мезофилла и регенерирует за счет энергии АТФ в ФЕП. СО2 вновь фиксируется РиБФ-карбоксилазой с образованием ФГК. Регенерация ФЕП требует энергии АТФ, поэтому нужно почти вдвое больше энергии, чем при С3-фотосинтезе.
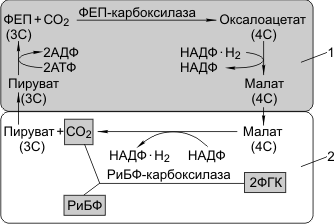
С4-фотосинтез:
1 — клетка мезофилла; 2 — клетка обкладки проводящего пучка.
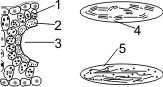
Строение С4-растений:
1 — наружный слой — клетки мезофилла; 2 — внутренний слой — клетки обкладки; 3 — «Кранц-анатомия»; 4, 5 — хлоропласты; 4 — многочисленные граны, крахмала мало; 5 — немногочисленные граны, крахмала много.
Значение фотосинтеза
Благодаря фотосинтезу, ежегодно из атмосферы поглощаются миллиарды тонн углекислого газа, выделяются миллиарды тонн кислорода; фотосинтез является основным источником образования органических веществ. Из кислорода образуется озоновый слой, защищающий живые организмы от коротковолновой ультрафиолетовой радиации.
При фотосинтезе зеленый лист использует лишь около 1% падающей на него солнечной энергии, продуктивность составляет около 1 г органического вещества на 1 м2 поверхности в час.
Источник: Лекции по общей биологии.